23 Oct Choanephora
Choanephora
Background
The genus Choanephora belongs to family Choanephoraceae in the order Mucorales (former Zygomycota). The genus was introduced by Currey (1873) for C. cunninghamii, to replace the generic name of his newly described species Cunninghamia infundibulifera, as Cunninghamia already existed as a genus of conifers. Because the specific epithet could not be retained, Choanephora cunninghamia remained invalid, based on the same type as Cunninghamia infundibulifera. The proper name Choanephora infundibulifera was validly published by Saccardo (1891), so the correct authorship of the species is “(Currey) Sacc.” It is also the type species of the genus. Choanephora was monographed by Hesseltine (1953), Milko and Beljakova (1970) and Kirk (1984). Currently, the genus is classified within the family Choanephoraceae which can be distinguished by the presence of a persistent sporangium wall that ruptures at preformed sutures. It is furthermore placed in the subfamily Choanephoroideae, which is characterized by the presence of apposed suspensors and smooth zygospores (Hoffmann et al. 2013).
Both species of the genus can grow as saprobes, but they frequently become plant pathogens causing various leaf and fruit rots and blights and are commonly reported from a wide range of plant hosts, including angiosperms (monocotyledons and dicotyledons) and gymnosperms (Farr and Rossman 2014). Their distribution is worldwide, however, disease development is more common in tropical and subtropical regions characterized by high temperatures and humidity. Choanephora cucurbitarum is the causal agent of fruit and blossom rot of various cucurbits, e.g. yellow crookneck squash (Kucharek and Simone 1983). This species is also known from crop plants such as green beans (McMillan 1972), garden peas (Oikawa et al. 1986), and okra (El-Sayed and El-Sajed 2013) and is reported as an agent of wet rot of Mesembryanthemum crystallinum in hydroponic greenhouse culture in Japan (Kagiwada et al. 2010). It is very common during rainy summers in the southeastern United States and globally in other regions with similar climates. Recently it was isolated also from cultivated Hyoscyamus muticus in Japan (Abdel-Motaal et al. 2010) and Withania in India (Saroj et al. 2012). Choanephora often attacks tissues that have been damaged mechanically by insects or otherwise; plants that are poorly adapted to a hot humid climate are particularly prone to infection by the genus. The general appearance of Choanephora rot is similar to that of blights caused by other Mucorales representatives. Signs of infections on fruits or leaves include water-soaked, necrotic lesions, which progress rapidly under wet conditions. As the fungus begins to produce spores, affected tissues become dark grey-brown and hairy. This specific appearance results from the tall sporangiophores that produce a cluster of brown, one-spored sporangia at their tips (Turkensteen 1979).
Species identification and numbers
Although more than ten species (and many varieties) have been described within this genus, only two species (viz. Choanephora infundibulifera and Choanephora cucurbitarum) were finally recognized in a monograph of the genus (Kirk 1984). These two species can be distinguished by the shape and ornamentation of indehiscent sporangia. C. cucurbitarum produces ellipsoid sporangia, which are usually distinctly longitudinally striate, whereas C. infundibulifera forms subglobose to obovoid sporangia with usually smooth or faint striate ornamentation. The remaining species were synonymized under these taxa (e.g. C. mandshurica is currently a synonym of C. cucurbitarum) or were moved to other genera (e.g. C. persicaria is a synonym of Gilbertella persicaria). Choanephora circinans with its two varieties (C. circinans var. indica and C. circinans var. prolifera) were moved by Kirk (1984) to Poitrasia. Poitrasia was established for those species belonging to the family Choanephoraceae that do not form dehiscent or indehiscent sporangia (Kirk 1984). Although Poitrasia is primarily a soil-borne genus, it has been isolated from Equisteum arvense (Rai 1990). Recent molecular studies confirmed the taxonomic position of Poitrasia proposed by Kirk (1984).
Molecular phylogeny
All Choanephora strains available in CBS culture collection (three strains of C. infundibulifera and five strains of C. cucurbitarum) have been sequenced for their ITS sequences and included in the molecular analysis by Walther et al. (2013). These studies showed that the universal fungal DNA barcoding marker–the ITS region (Schoch et al. 2012)–is sufficient for Choanephora species identification (Table, Fig. ). A multigene phylogenetic analysis including representatives of this genus was performed by Hoffmann et al. (2013).
Table Choanephora. Details of the isolates used in the phylogenetic tree
| Species | Isolate | Host | GenBank no |
| Choanephora infundibilifera | CBS 153.51 | – | JN206236 |
| C. infundibilifera | CBS 155.51 | – | JN206237 |
| C. infundibilifera | CBS 155.58 | – | JN206238 |
| C. cucurbitarum | CBS 445.72 | – | JN206234 |
| C. cucurbitarum | CBS 178.76 | Dead insect | JN206235 |
| C. cucurbitarum | CBS 674.93 | – | JN206233 |
| C. cucurbitarum | CBS 120.25 | – | JN206231 |
| C. cucurbitarum | CBS 150.51 | – | JN206232 |
| Poitrasia circinans | CBS 153.58* | Soil | JN206239 |
| P. circinans | CBS 647.70 | Soil | JN206240 |
Ex-type (ex-epitype) strains are bolded and marked with an * and voucher strains are bolded
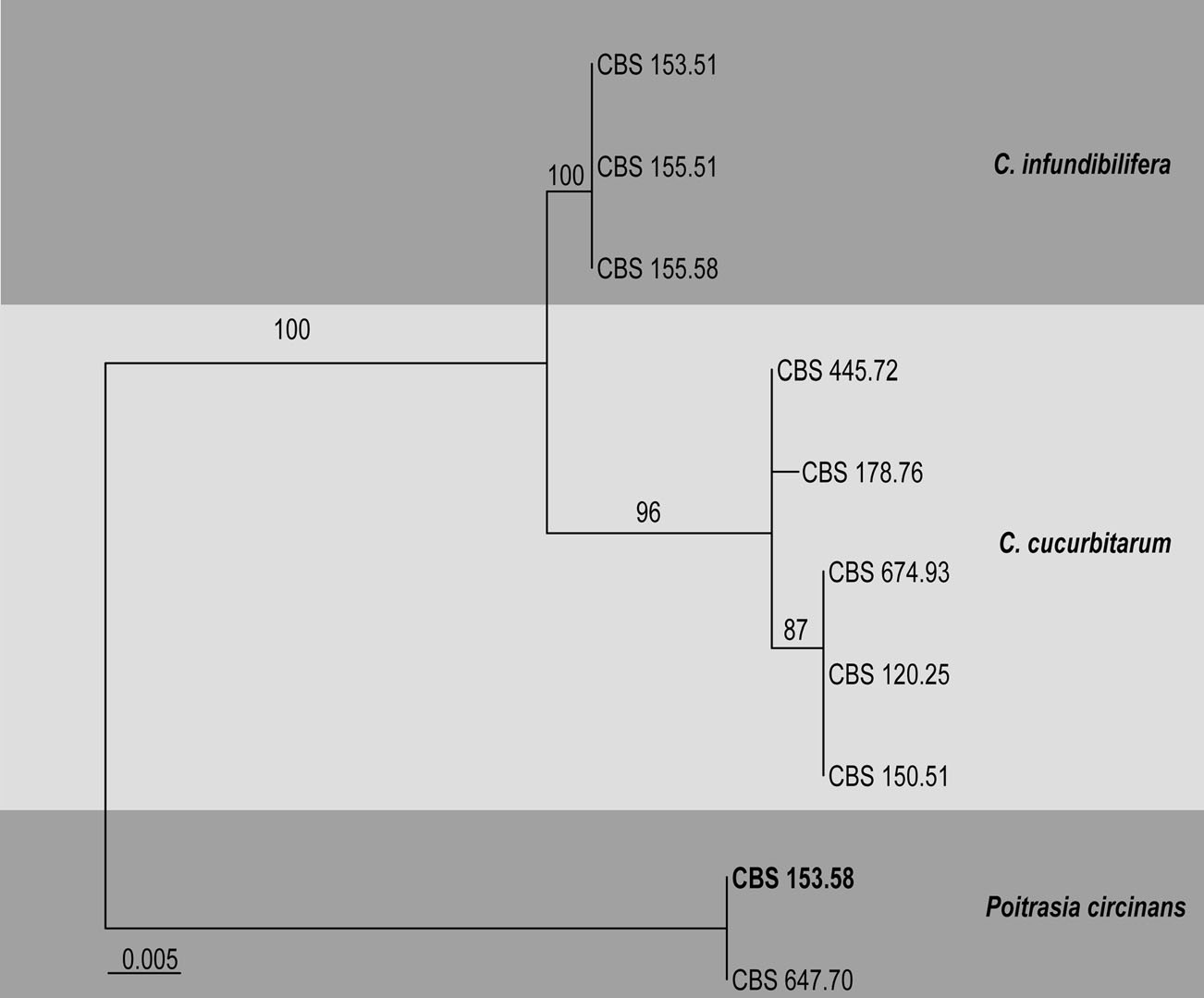
Fig. Phylogramgenerated from Maximum likelihood analysis based on ITS sequenced data of Choanephora. Bootstrap support values greater than 50 % are indicated above the nodes. The ex-type (ex-epitype) and voucher strains are in bold
Recommended genetic markers
- The internal transcribed spacer (ITS)–generic and species level
- The large and small subunits (LSU and SSU) of nrDNA– placement within the Mucorales order, higher-level phylogeny
- The partial actin gene (ACT) and the partial translation elongation factor 1-alpha gene (TEF)–higher-level phylogeny
No Comments